| pA2 online © Copyright 2004 The British Pharmacological Society |
044P
University of Newcastle Winter Meeting December 2004 |
Cannabinoids prevent β-amyloid-induced microglial activation and neurotoxicity B.G. Ramírez1, C.Blázquez2, T.Gómez del Pulgar2, M.Guzmán2 & M.L. de Ceballos1. 1Neurodegeneration Group, Cajal Institute, CSIC, 28002 Madrid, Spain.2Department of Biochemistry and Molecular Biology I, School of Biology, Complutense University, 28040 Madrid, Spain. |
|
Alzheimer’s disease (AD) brains develop plaques in which ß-amyloid (ßA) deposits and activated microglia exist. Microglia express CB1 and CB2 receptors and cannabinoids reduce cytokine production in microglial cell cultures. Cannabinoids (cbs) are neuroprotective agents against excitoxicity in vitro and acute brain damage in vivo. These prompted us to study the effects of cbs on βA-induced neurotoxicity and microglial activation in vitro.
Microglial cells were obtained by shaking primary mixed glial cultures from neonatal rat cortex, after being cultured for 3 weeks, and 3 x 104 cells cells were seeded onto 96-well plates in DMEM/Ham’s F12 /0.5 % FCS. Mitochondrial activity was assessed by the MTT assay. TNF-α was measured by a commercial ELISA. Neurons from E17 fetal rat cortex 25 x 104 cells were seeded onto 24 well plates in Neurobasal containing B27 additives After 4 days in culture they were switched to MEM with N2 additives, and treatments (DIV 6) were performed in MEM. For co-cultures, 15 x 104 microglial cells were seeded in inserts, at 24 h were treated with βA1-40 (sol:soluble or fib:fibrillar, after aging at 37º C for 24 h; Neosystem, France) for 4 h to avoid direct toxicity on neurons, the medium was aspirated and placed over neuron cultures for a further 20 h period. Neurons and microglia were counted following Coomassie staining. Results are mean±s.e.m. of at least 4 independent experiments done in triplicate.
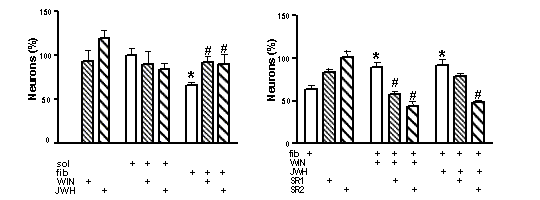
Fig 1. Cbs prevent βA-induced microglial neurotoxicity. Reversal by cb antagonists: left panel *p<0.05 versus sol-treated; #p<0.05, vs fib-treated (ANOVA with Bonferroni post hoc test); right panel *p<0.05 versus cultures treated with fib alone; #p<0.05, vs fib plus the CB agonist.
HU-210 (100 nM), prevented ßA-induced (500 nM) microglial activation which included morphological changes, increased mitochondrial activity and TNF-α release. WIN55,212-2 (WIN) and JWH-133 (JWH), a CB2 selective agonist, mimicked the latter effect. Both WIN and JWH prevented microglia-mediated neurotoxicity following ßA treatment (Fig. 1) . The neuroprotective effect of WIN on ßA neurotoxicity in co-cultures was blocked by the selective antagonists SR141716 (SR1) and SR144528 (SR2), while the effect of JWH was only counteracted by the latter (Fig. 1).
Since cannabinoids combine both antiinflammatory and neuroprotective actions, our findings may set the basis for their use as a therapeutic approach for AD.