
Contraction induced by noradrenaline or serotonin is increased in the rat superficial epigastric artery during lactation The superficial epigastric arteries (SEAs) supply the inguinal fat pads and mammary glands in rats. Although these arteries are important in supplying blood to skin flaps in rat models for plastic surgery (e.g. Lay, et al., 2003; O’Neill, et al., 2006) , little is known about the pharmacology of this vessel, or changes which may occur at peak lactation when the demands on the mammary gland are greatest. The aim this study therefore was to investigate the response of rat SEAs to two classical vasoactive compounds outwith, and during peak lactation. Female Sprague-Dawley rats (180-350g) which were either a) non-pregnant and non-lactating, or b) between days 11-18 of lactation were used. The animals were killed by giving a lethal dose of sodium pentobarbitone (Sagatal; 120 µg/g body weight i.p.). The superficial epigastric arteries (SEA) were dissected, cleaned of connective tissue, and separated from the parallel vein and nerve. Lengths of 1.5 – 2.0 mm were cut and mounted in a conventional Mulvany Myograph (Danish Myotech, Aarhus, Denmark) for recordings in a HEPES-buffered Tyrode solution (pH 7.4) gassed at 37°C with 100% O2. The Tyrode solution also contained propranolol (1 µM), cocaine (3 µM), and corticosterone (15 µM). After equilibrating for at least 45 min, the arteries were stretched to a force of 2.5 mN (previously determined to be the optimal passive force), and allowed to re-equilibrate at that tension. Viability of the preparations was determined by exposing to a high potassium solution (80 mM final concentration) until two consecutive comparable contractions were obtained; no steps were taken to denude the preparation of endothelium. Cumulative concentration response curves (CRCs) were obtained by adding noradrenaline (NA) or serotonin (5-hydroxy-tryptamine; 5-HT) at final concentrations of 10-7-10-4 or 10-9-10-5 M respectively. Data are mean ± sem; n = number of animals; Student’s t test was used to test differences. Rat SEAs responded to both NA and 5-HT CRCs in a dose dependent manner (Figure 1A); the pEC50 for NA was significantly smaller (p < 0.001) than that for 5-HT (Figure 1B).
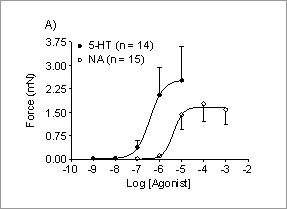 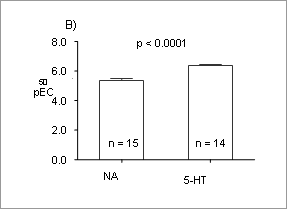
Figure 1. A) CRCs for control SEA using NA or 5-HT, and B) pEC50 for NA or 5-HT in control SEA.
The maximal effect (Emax) obtained with both NA or 5-HT CRCs was greater in lactating animals than controls: 4.97 ± 1.18 mN (n=18) vs. 1.87 ± 0.60 mN (n = 15) for NA (p < 0.05) and 6.59 ± 1.12 mN (n=16) vs. 2.68 ± 1.15 mN (n=14) for 5-HT (p < 0.02). There were no significant differences when comparing the Emax values for NA with 5-HT either in the control or lactating animals. The pEC50 was significantly less in control animals compared with lactating rats for both agonists: NA; 5.37 ± 0.08 (n=15) vs. 5.67 ± 0.09 (n = 18; p < 0.005), and 5-HT; 6.39 ± 0.04 (n=14) vs. 6.70 ± 0.06 (n = 16; p < 0.0005) respectively. We conclude that during peak lactation, the rat SEA is more responsive to the agonists NA or 5-HT than when not lactating.
Lay, I. S., et al., (2003). J. Surg. Res., 115, 279-85. |
|
