

CAATs at the mouse choroid plexus Introduction - Arginine, an essential precursor in the synthesis of nitric oxide (NO), can not be synthesised de novo in endothelial cells (Am J Physiol Heart Circ Physiol, 265 (1993) H1965-H1971). NO is synthesised in the capillary endothelium (such as that found at the blood-brain interface) by endothelial nitric oxide synthase (eNOS, Neurosci Lett, 239 (1997) 109-112); where it has explicit roles in cellular signalling, vasodilatation and the immune response (Nat Rev Drug Discov, 1 (2002) 939-950). NO synthesis is known to be inhibited by endogenous asymmetric dimethylarginine (ADMA); believed to be due to the inability of eNOS to use ADMA as a substrate (Lancet, 339 (1992) 572-575). Changes in the ratio of L-arginine:ADMA have been implicated in many clinical conditions (Nat Med 13 (2007) 198-203). The transport kinetics of both these cationic amino acids into the brain and highly vascularised circumventricular organs (e.g. choroid plexus) can be expected to provide valuable insights into the molecular pathology of conditions such as stroke. Little is known about the cationic amino acid transporters (CAATs) at the choroid plexus, although L-arginine transport at the rat choroid plexus is known to be saturable and sensitive to inhibition by another methylated arginine, L-NMMA (J Neurochem, 65 (1995) 68-74). The sensitivity to ADMA of the CAAT(s) responsible for arginine uptake is not yet known and is the object of this study.
Methods - The in situ brain perfusion technique was used to investigate arginine uptake at the choroid plexus in Balb/c mice (J. Pharm. Exp. Ther. 329 (2009) 967–977.). [3H]-labelled arginine and a vascular /extracellular space marker ([14C]sucrose) were transcardially perfused for 10 minutes in the absence or presence of either un-labelled arginine or un-labelled ADMA at 100µM.
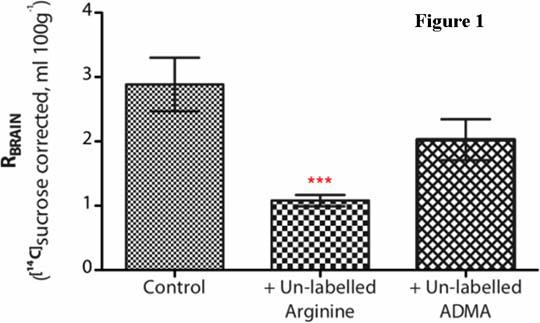
Results - Preliminary results ([14C]sucrose corrected) are shown in figure 1. [3H]arginine uptake at the choroid plexus is significantly reduced in the presence of un-labelled arginine, but not un-labelled ADMA. Graph shows Mean±SEM; n=5-7. ***P < 0.001, one-way ANOVA with Dunnett’s post-hoc test. 
Conclusions - L-arginine transport at the murine choroid plexus is saturable and is not significantly sensitive to ADMA. It remains to be elucidated which CAAT is responsible for arginine transport in this mouse tissue. Further work is on-going.
Acknowledgements - BBSRC 4 yr PhD programme. |
|