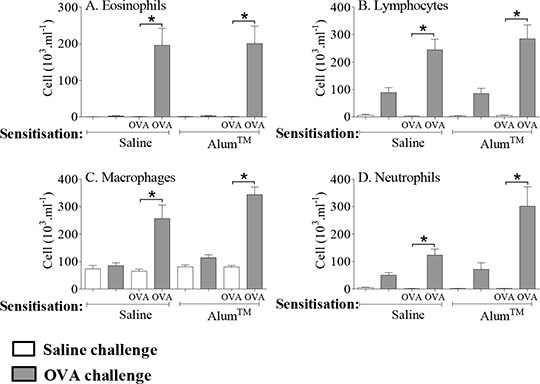
Is AlumTM Required To Generate An Allergic Asthmatic Phenotype In The Ovalbumin Model? The commonly used allergen-driven models of asthma all have caveats. Notably, the clinical relevance of the ovalbumin (OVA)-driven model may be limited by the use of systemic sensitisation with an adjuvant, typically AlumTM. The present study aims to determine whether the presence of AlumTM during sensitisation is necessary for an OVA-induced allergic response in this model. Male C57Bl/6 mice (18-20 grams) were sensitised with saline or OVA (10µg per mouse, in 100µl, i.p.), made up in the absence or presence of AlumTM, on days 0 and 14, and subsequently challenged intranasally with saline or OVA (50µg per mouse, in 50µl) on days 24-26. Approximately 72 hours after final challenge, airway response to inhaled saline and 5-HT (1-10 mg.ml-1) was assessed (whole body plethysmography (WBP); Penh). Total and OVA-specific IgE were measured in plasma by ELISA, and inflammatory cellular infiltrate was assessed in the bronchoalveolar lavage (BAL) fluid. In a similar model, OVA-induced late asthmatic response (LAR) was also measured immediately after challenge. OVA challenge in OVA-sensitised mice resulted in a significant increase in total and OVA-specific IgE, both in the absence and presence of AlumTM (n=7-9/group *=p<0.05, Mann-Whitney U-test vs OVA-sensitised/saline challenged controls) (Table 1). OVA challenge resulted in an increase in eosinophils, lymphocytes, macrophages, and neutrophils in mice sensitised with OVA, both in the absence, and presence of AlumTM (Figure 1). Table 1:
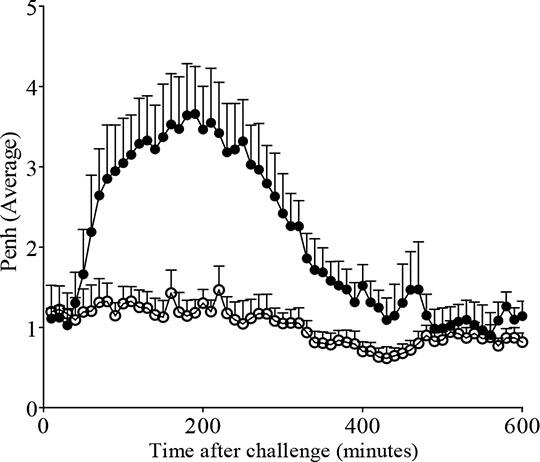
Finally, airway hyperresponsiveness to 5-HT was observed in OVA- sensitised and –challenged mice, irrespective of the use of AlumTM. Penh area under curve values (3 mg.ml-1 5-HT) were: OVA/saline 1055±81.96, OVA/OVA 1566*±130.1, AlumOVA/saline 872.1±66.65, AlumOVA/OVA 1474*±67.85 (*=p<0.05 vs OVA/saline or AlumOVA/saline, Mann-Witney U-test, 8-9/group). OVA-induced LAR, a key symptom of allergic asthma in man, was only observed in mice which were sensitised to OVA in the presence of AlumTM (Figure 2). Contrary to current dogma and recent publications (Kool et al. 2008), the use of a non-physiological adjuvant was not required to induce allergic responses in this model. However, AlumTM is required to observe the late asthmatic response. This provokes the question as to what is driving the LAR, as this response has been linked with increases in allergen-directed IgE (Fahy et al. 1997). 
Figure 1: Role of AlumTM during sensitisation in the response to OVA. Data expressed as mean cell number (103.ml-1) + SEM (n=7-9/group). *=p<0.05 vs saline- or AlumTM-OVA sensitised/saline challenged controls, Mann-Whitney U-test. 
Figure 2: The role of AlumTM in the OVA-driven LAR. Immediately after challenge (2% OVA in saline, i.t), LAR was recorded (WBP). Data expressed as mean Penh average + SEM for (n= 6-8/group).
Fahy, J. V.,et al. (1997) AJRRCM 155: 1828–1834. Kool, M., et al JEM 205: 869–882.
|