Print version
Search Pub Med

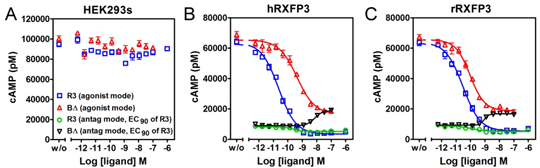
R3(BΔ23-27)R/I5 is a partial RXFP3 agonist – investigation of in vivo and in vitro pharmacology BACKGROUND AND PURPOSE RXFP3 is a G-protein coupled receptor mainly expressed in the brain, reported to be involved in appetite regulation and stress response. Previous studies in rats have shown that the chimeric peptide R3(BΔ23-27)R/I5 suppresses food intake stimulated by an RXFP3 agonist (1). We wanted to investigate if R3(BΔ23-27)R/I5 on its own is able to antagonize the basal tone of the relaxin-3/RXFP3 system, with potential effects on food intake. We therefore characterized the pharmacology of R3(BΔ23-27)R/I5 in vivo and in vitro. EXPERIMENTAL APPROACH Food intake was measured in male WistarHan rats intracerebroventricularly (ICV) injected with R3(BΔ23-27)R/I5 or the endogenous RXFP3 agonist relaxin-3. R3(BΔ23-27)R/I5 was subsequently characterized in vitro using [35S]-GTPγS binding and cAMP assays, both assessing Gα i-protein mediated signaling, and label-free dynamic mass redistribution (DMR) assays capturing the integrated cell response. KEY RESULTS ICV injection of relaxin-3 stimulated food intake in satiated lean rats in line with previous results (2). R3(BΔ23-27)R/I5 alone, dosed into diet induced obese (DIO) rats four hours before the beginning of the dark phase when feeding drive is high, did not have any significant effect on food intake. In vitro, R3(BΔ23-27)R/I5 showed potent partial agonist activity in [35S]-GTPγS binding, cAMP and DMR functional assays. Figure 1. Characterization of relaxin-3 and R3(BΔ23-27)R/I5 in cAMP assays.

HEK293s (A), HEK293s-hRXFP3 (B), HEK293s-rRXFP3 (C) cells were co-treated with 10 µM forskolin (to reveal Gi-dependent signalling) and concentrations of relaxin-3 (R3) or R3(BΔ23-27)R/I5 (BΔ) as indicated. Concentration responses in (B) and (C) were run either in the absence (agonist mode) or presence of 0.8nM relaxin-3 (R3@EC90, antagonist mode). For relaxin-3, pEC50s were 10.59 ± 0.08 (hRXFP3) and 10.44 ± 0.11 (rRXFP). For R3(BΔ23-27)R/I5, pEC50s and Emax (compared to relaxin-3) were 9.41 ± 0.16, 83 ± 5 % (hRXFP3) and 10.05 ± 0.14, 78 ± 3 % (rRXFP3). CONCLUSIONS AND IMPLICATIONS R3(BΔ23-27)R/I5 is a partial RXFP3 agonist in vitro. Hence, it is not an ideal tool to address if antagonizing relaxin-3/RXFP3 basal tone is a therapeutically relevant mechanism to regulate food intake and body weight in vivo, and identification of more appropriate tools are required. (1) Kuei C et al, J Biol Chem 282:25425, 2007 (2) McGowan BMC et al, Endocrinol 146:3295, 2005
|